Опубликовано:
Клиническая дерматология и венерология 1, 2012Аспартат протеазы грибов рода Candida – потенциальная мишень для антивирулентной терапии
И.В. Чеботарь1
ГОУ ВПО Нижегородская государственная медицинская академия Минздравсоцразвития РФ, Нижний НовгородAspartate proteases of Candida fungi – a potential target for anti-virulent therapy
I.V. Chebotar’
State budgetary educational institution of higher professional education Nizhni Novgorod State Medical Academy, Ministry of Health and Social Development of the Russian Federation, Nizhni NovgorodКлючевые слова: кандидоз, грибы рода Candida, фактор патогенности, вирулентность, секретируемые аспартат протеазы, антивирулентная терапия.
Key words: candidiasis, fungi of the genus Candida, pathogenicity factor, virulence, secretory aspartate proteases, antivirulent therapy.Наиболее существенное значение в патологии человека имеют грибы рода Candida, которые относятся к оппортунистическим инфекциям и являются актуальной проблемой современной медицины. Опасность возникновения кандидозов и тяжесть их течения возрастает при длительной антибактериальной химиотерапии, а также при разнообразных иммунодефицитных состояниях [1]. Грибы рода Candida реализуют свой болезнетворный потенциал через набор факторов патогенности, к числу которых относятся адгезины (семейство адгезинов Als, Hwp1, Int1), гифы, внеклеточные энзимы-гидролазы — фосфолипаза Б, липазы, аспартат протеазы [22].
Важнейшим фактором патогенности, обеспечивающим деструкцию тканей человека, являются аспартат протеазы [16], которые, концентрируясь на терминальных окончаниях гиф, обеспечивают непосредственную инвазию кандид в ткани человека. Доктором B. Hube отмечено, что патогенность кандид является мультифакторной, а вирулентность определяется аспартат протеазами [16]. Следовательно, возможность нейтрализации аспартат протеаз может стать эффективным инструментом антивирулентной терапии при кандидозах.
Цель работы — структурно-функциональная характеристика аспартат протеаз дрожжеподобных грибов рода Candida и поиск возможных путей фармакологической нейтрализации их активности.
Аспартат протеазы (синоним — аспартильные протеазы) — семейство ферментов-протеаз, в составе активного центра которого имеются остатки аспарагиновой кислоты (Asp) для катализа соответствующих пептидных субстратов. В англоязычной литературе они обозначаются как «aspartic proteases» и имеют много синонимов: «aspartic proteinases», «aspartic endopeptidases», «aspartate proteases», «aspartictype endopeptidases», «aspartyl proteinases», реже — «acid proteases». В русскоязычных научных текстах часто используется термин «аспартаткислые протеазы» для того, чтобы подчеркнуть оптимальную активность этих ферментов при кислых значениях pH.
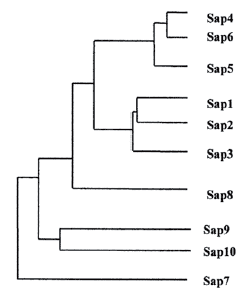
В общей биохимической классификации белков по своим молекулярно-функциональным характеристикам они относятся к классу ферментов-гидролаз, гидролизующих пептидные связи в белках, т.е. принадлежат к подклассу ферментов-протеаз. Изученные аспартат протеазы дрожжеподобных грибов являются эндопептидазами, т.е. ферментами, гидролизирующими внутренние, α-пептидные связи в полипептидных цепях (этим они отличаются от амино- и карбоксипротеаз, отщепляющих от полипептидной цепи концевые аминокислоты). Согласно классификации UniProtKB (http://www.uniprot.org), группа EC 3.4.23 «Aspartic endopeptidases» включает более 40 видов аспартат протеаз, выделенных от представителей разных таксономических групп — грибов, позвоночных (в том числе млекопитающих), растений, вирусов [27]. Типовой аспартат протеазой кандид является кандидапепсин (candidapepsin), который относится к группе секретируемых аспартат протеаз (САП; англ. акроним — Sap). В настоящее время установлено, что важнейший в патогенетическом отношении представитель кандид — Candida albicans секретирует 10 типов САП. Соответственно, доказано существование 10 генов C. albicans, кодирующих белки семейства САП [26]. На рис. 1. представлена дендрограмма, созданная J. Naglik и соавт. [22]: она демонстрирует наличие трех разных групп изоферментов внутри семейства САП. В целом гомология между разными формами этого фермента составляет от 98% (между САП 4 и САП 6) до 20-27% (между САП 7 и другими представителями этого семейства).
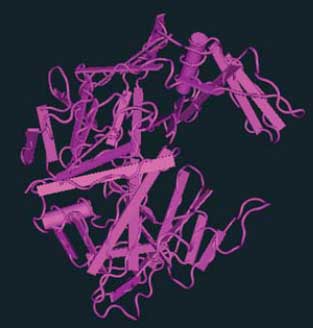
Рис. 1. Дендрограмма изоферментов группы аспартат протеаз Сandida albicans [22].Полная последовательность строения аспартат протеазы C. albicans была изучена в середине 90-х годов XX века [2]. В этой же работе была построена трехмерная модель молекулы САП. В настоящее время можно легко познакомиться как с аминокислотными последовательностями, так и с трехмерной организацией разных вариантов семейства САП, используя ресурсы базы данных Structure (Национальный центр биотехнологической информации, США). Например, на этом сайте представлен великолепный образец реконструкции 3D-структуры (программа Cn3D 4,3) кандидапепсина 1 или САП 1 (рис. 2), построенный на основе данных C. Borelli и соавт. [6].
Рис. 2. Виртуальная 3D-модель кандидапепсина 1 (САП 1), построенная c помощью программы Cn3D 4,3 (Национальный центр биотехнологической информации, США) на основании данных C. Borelli и соавт. [6].Классическим ингибитором аспартат протеаз является пепстатин.
Молекулярная масса аспартат протеаз кандид составляет около 40 кД и варьирует от 37 кД у САП 5 до 49 у САП 2 [2]. Оптимальная активность разных изоформ САП варьирует в пределах значений рН от 3,2—3,5 (САП 2 и САП 3), 3,2—4,5 (САП 1) до 5,0 (САП 4, САП 5, САП 6) [2].
Ферменты семейства САП продуцируются многими видами рода Candida: они идентифицированы у С. albicans, C. tropicalis, C. parapsilosis, C. lusitaniae, C. dubliniensis [22].
Сценарий кандидоза определяется субстратами, которые могут стать мишенями для аспартат протеаз. Субстратами для САП могут служить разнообразные простые и сложные белки человека. Среди них — молекулы, защищающие слизистые оболочки человека, например, муцин или секреторный IgA [8, 15]. Доказано, что САП может вызывать прямую деградацию целой группы неспецифических факторов защиты слизистых оболочек — лактоферрина, лактопероксидазы, катепсина D и факторов комплемента [14, 16, 17]. Кроме того, САП могут гидролизировать такие белки, как цистатин А, являющийся ингибитором цистеинпротеазы в клетках человеческого эпидермиса [28]. Инвазия непосредственно в эпителиоциты также осуществляется за счет САП. Активность изоферментов САП в процессе инвазии в ткани может существенно различаться. На модели человеческих кератиноцитов показано, что экспрессия семейства генов САП наблюдалась в начале инвазии в следующем порядке: САП 1 и САП 2>САП 8>САП 6>САП 3; глубокая инвазия — в большей степени САП 8 [26]. Пепстатин А ингибировал процесс повреждения тканей; результаты наблюдения за САП-дефицитными мутантами кандид также свидетельствовали о корреляции между активностью САП и повреждением тканей [26]. Аспартатпротеазы могут оказывать на ткани непрямой повреждающий эффект через активацию воспаления. САП способны вызывать стимуляцию флогогенеза, обеспечивая трансформацию прекурсора интерлейкина-1β (ИЛ-1β) в полноценный (ИЛ-1β), который является цитокином с выраженной провоспалительной активностью [5]. Еще один флогогенный эффект, связанный с САП, был изучен на модели человеческого вагинального эпителия [25]. В этой работе была доказана САП-зависимая стимуляция выработки ИЛ (ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-10), гранулоцитарно-макрофагального колоний-стимули рующего фактора, γ-интерферона (относительно экспрессии цитокинов в неинфицированной ткани). Важнейший плазменный фактор — α2-макроглобулин человека также подвергается САП-опосредованной деградации [24].
Все перечисленные случаи вскрывают механизмы, с помощью которых кандиды реализуют свой инвазивный потенциал. Неоспоримые доказательства ведущей роли САП в развитии кандидоза получены в многочисленных опытах на животных [22]. Особого внимания заслуживают эксперименты по моделированию местного или диссеминированного кандидоза, в которых активность аспартат протеаз подавлялась ингибиторами (пепстатин); в этих случаях развития патологического процесса не отмечено [13, 22]. Подобные результаты наблюдались при попытке моделирования кандидозного вагинита у крыс с использованием анти-САП-антител, которые оказывали выраженный протективный эффект, тормозя развитие инфекционного процесса [12]. Все эти факты служат прямыми доказательствами того, что аспартат протеазам принадлежит главная роль в патогенезе разных форм кандидоза. Существуют и косвенные доказательства, к которым относятся корреляция между вирулентностью кандид и уровнем продукции САП in vitro, повышение уровня экспрессии генов САП при кандидозной инфекции (in vivo), корреляция между САП-продукцией и иммунным ответом хозяина, экспрессией САП и другими факторами патогенности кандид, авирулентность многих штаммов-мутантов кандид, не способных секретировать САП [22].
Однако требуется существенное уточнение. Аспартатпротеазы – ведущий, но не единственный фактор инвазии кандид. В работе U. Lermann и J. Morschhauser доказано, что филаменты мутантов C. albicans, не способных к секреции аспартат протеаз, могут опосредовать вирулентность кандид. Штаммы-мутанты, дефектные по САП и филаментам efg1D, были авирулентны [19].
Говоря о патогенности, нужно вспомнить о возможности кандид формировать особые микробные сообщества — биопленки [10]. Эту способность в настоящее время можно расценивать как фактор патогенности: внутри биопленок микробы приобретают выраженную устойчивость к эффекторам иммунной защиты и противомикробным препаратам. При росте в форме биопленок C. albicans проявляла более выраженную секрецию аспартат протеаз по сравнению с существованием в планктонных формах. Авторы [20], наблюдавшие этот феномен, склонны расценивать его как один из механизмов повышения вирулентности кандид в составе биопленок.
Специфическая инактивация аспартат протеаз возможна при использовании трех подходов: 1) классического метода нейтрализации факторов патогенности на основе антител; 2) создании химиопрепаратов, специфично ингибирующих САП за счет прямого связывания; 3) поиска препаратов, негативно регулирующих экспрессию САП (на уровне генов или посттрансляционных и/или посттранскрипционных событий).
Возможность реализации первого подхода проверена лишь в экспериментальных работах. Нами уже упоминалась работа, в которой показана протективная роль антител к САП при экспериментальных кандидозных вагинитах [12]. К сожалению, за прошедшее с момента публикации время никому не удалось реализовать изложенную в ней идею — препараты на основе анти-САП-антител в клинической практике так и не появились. Это может свидетельствовать о недостатках подобного подхода к антивирулентной терапии кандидоза.
В экспериментах in vitro и in vivo показана возможность прямой специфической блокады САП пепстатином, при этом активность кандидозного процесса снижалась [22]. Однако пепстатин не является специфичным для грибковых САП и блокирует активность аспартат протеаз млекопитающих. С этих позиций широкое применение пепстатина для лечения кандидозов является сомнительным.
Первые научно обоснованные теории, предлагающие пути создания фармацевтических препаратов, прямо инактивирующих САП, были предложены в 1996 г. С. Abad-Zapatero и соавт. [2]. В этой работе была создана 3D-модель кандидапепсина и намечены первые уязвимые участки молекулы, воздействие на которые должно вызвать «драматичную альтерацию» фермента. Более того, было доказано, что эти области специфичны для кандид и не встречаются в составе аспартат протеаз млекопитающих — ренина, пепсина, гастрина. Такая автономность, по справедливому мнению авторов, позволяет создать высокоспецифичные ингибиторы главных факторов вирулентности кандид — ферментов семейства САП.
В конце ХХ века появились антиретровирусные препараты (для лечения ВИЧ-инфекции), действие которых было направлено на ингибирование ВИЧспецифических ферментов — ревертазы (обратной транскриптазы), интегразы и ретровирусной аспартат протеазы. Практически сразу же после появления препаратов-ингибиторов ретровирусной аспартат протеазы (ритонавир, саквинавир, индинавир) были предприняты успешные попытки использовать их для лечения экспериментальных кандидозов, вызванных C. albicans [7, 18]. Антикандидозный эффект индинавира и ритонавира был сравним с эффектом флуконазола [7]. В работе I. Pichova и соавт. [23] активность ритонавира и саквинавира показана в отношении кандидозов, вызванных разными представителями рода Candida – С. albicans, C. tropicalis, C. parapsilosis, C. lusitaniae.
Серия работ была посвящена исследованию влияния противогрибковых химиопрепаратов на секрецию кандидами аспартат протеаз. Результаты оказались неожиданными. Субингибирующие концентрации флуцитозина, флуконазола, итраконазола, миконазола и каспофунгина вызывали активацию САП, хотя клеточный рост при обработке итраконазолом, миконазолом, флуцитозином и каспофунгином замедлялся [9]. Даже резистентный к азолам и флуцитозину штамм C. albicans демонстрировал повышение секреции аспартат-протеаз при обработке флуконазолом [9]. Топически активация САП 4 и САП 6 под воздействием флуконазола и флуцитозина наблюдалась в расту щих клетках [4]. Удивительным оказалось то, что другой представитель семейства азолов — фентиконазол вызывал стойкое ингибирование протеиназной активности C. albicans даже в концентрациях 0,01 и 1—10 мкг/мл, 5-флуороцитозин ингибировал активность САП в концентрации 1—10 мкг/мл [3]. Подобные результаты получены и в другой работе, в которой фентиконазол в отличие от эконазола тоже вызывал сильное подавление активности САП [11]. В обоих случаях авторы обсуждают полученный эффект с позиции непрямого взаимодействия фентиконазола и протеаз; наиболее вероятное объяснение ингибирования САП связано с воздействием на общие механизмы синтеза белка. Попытка непрямого подавления протеазной секреции C. albicans была сделана в работе S. Milewski и соавт. [21]. Авторы использовали антигрибковый агент L-lysyl-L-norvalyl-[N3-(4-methoxyfumaroyl)]L-2,3-diamino pro panoic acid (Lys-NvaFMDP), который в минимальных концентрациях угнетал секрецию САП. При этом специфичность ингибирования была выше, чем при использовании фентиконазола.
Анализ большого объема научной информации, посвященной исследованиям аспартат протеаз у грибов рода Candida, позволяет рассматривать семейство САП в качестве системы, определяющей вирулентность кандид. Подавление вирулентности за счет специфического ингибирования аспартат протеаз подтверждает тезис о том, что аспартат протеазы могут стать основной мишенью для антивирулентной терапии кандидоза. Можно с уверенностью заключить, что современные достижения молекулярного моделирования позволят создать новые высокоспецифичные препараты для блокады секретируемых аспартат протеаз.
Литература
- Сергеев А.Ю., Сергеев Ю.В. Кандидоз. Природа инфекции, механизмы агрессии и защиты, лабораторная диагностика, клиника и лечение. М: Триада-X 2001; 472.
- Abad-Zapatero С., Goldman R., Muchmore S.W. et al. Structure of a secreted aspartic protease from C. albicans complexed with a potent inhibitor: Implications for the design of antifungal agents. Protein Sci 1996; 5: 4: 640—652.
- Angiolella L., De Bernardis F., Bromuro C. et al. The effect of antimycotics on secretory acid proteinase of Candida albicans. J Chemother 1990; 2: 1: 55—61.
- Barelle C.J., Duncan V.M.S., Brown A.J.P. et al. Azole antifungals induce up-regulation of SAP4, SAP5 and SAP6 secreted proteinase genes in filamentous Candida albicans cells in vitro and in vivo. J Antimicrob Chemother 2008; 61: 2: 315—322.
- Beausejour A., Grenier D., Goulet J. P., Deslauriers N. Proteolytic activation of the interleukin-1 beta precursor by Candida albicans. Infect Immun 1998; 66: 2: 676—681.
- Borelli C., Ruge E., Lee J.H. et al. X-ray structures of Sap1 and Sap5: structural comparison of the secreted aspartic proteinases from Candida albicans. Proteins 2008; 72: 4: 1308—1319.
- Cassone A., De Bernardis F., Torosantucci A. et al. In vitro and in vivo anticandidal activity of human immunodeficiency virus protease inhibitors. J Infect Dis 1999; 180: 2:453—488.
- Colina A.R., Aumont F., Deslauriers N. et al. Evidence for degradation of gastrointestinal mucin by Candida albicans secretory aspartyl proteinase. Infect Immun 1996; 64: 11: 4514—4519.
- Copping V.M., Barelle C.J., Hube B. et al. Exposure of Candida albicans to antifungal agents affects expression of SAP2 and SAP9 secreted proteinase genes. J Antimic Chemother 2005; 55: 5: 645—654.
- Costerton J.W., Stewart P.S., Greenberg E.P. Bacterial biofilms: a common cause of persistent infections. Science 1999; 284: 1318— 1322.
- De Bernardis F., Cassone A. Comparison of the effects of fenticonazole and econazole on the aspartic proteinase secreted by Candida albicans. Contracept Fertil Sex 1996; 24: 2: 163—165.
- De Bernardis F., Boccanera M., Adriani D. et al. Protective role of antimannan and anti-aspartyl proteinase antibodies in an experimental model of Candida albicans vaginitis in rats. Infect Immun 1997; 65: 8: 3399—3405.
- Fallon K., Bausch K., Noonan J. et al. Role of aspartic proteases in disseminated Candida albicans infection in mict. Infect Immun 1997; 65: 2: 551—556.
- Germaine G.R., Tellefson L.M. Effect of pH and human saliva on protease production by Candida albicans. Infect Immun 1981; 31: 323—326.
- Goldman R.C., Frost D.J., Capobianco J.O. et al. Antifungal drug targets: Candida secreted aspartyl protease and fungal wall beta-glucan synthesis. Infect Agents Dis 1995; 4: 4: 228—247.
- Hube B. Candida albicans secreted aspartyl proteinases. Curr Top Med Mycol 1996; 7: 1: 55—69.
- Kaminishi H., Miyaguchi H., Tamaki T. et al. Degradation of humoral host defense by Candida albicans proteinase. Infect Immun 1995; 63: 3: 984—988.
| Апрель 2012 г. |