Опубликовано:
Гинекология, том 14, №2«Быстрый эффект» клималанина (бета-аланина) при приливах: сравнительное исследование взаимодействий бета-аланина, таурина и глицина с глициновыми рецепторами
Торшин И.Ю.1, Громова О.А.1,2, Лиманова О.А.1,2
1Российский сателлитный центр института микроэлементов ЮНЕСКО, Москва
2Кафедра фармакологии и клинической фармакологии ГБОУ ВПО Ивановская государственная медицинская академия Минздравсоцразвития РФАбстракт
Бета-аланин – неканоническая аминокислота, содержащаяся в препарате клималанин, использующаяся в терапии «приливов» и позволяющая быстро, в течение считанных минут, купировать возникающий сосудистый пароксизм. В настоящей работе впервые проведено сравнительное биофизическое моделирование взаимодействий бета-аланина с пространственной структурой глициновых рецепторов.
Показано, что энергия связывания рецептором бета-аланина превышает энергию связывания самого глицина, что соответствует пролонгированному (по сравнению с глицином) воздействию препаратов бета-аланина на глициновые рецепторы, оказывая нормализующее действие на терморегуляцию и вазомоторику.
Ключевые слова: приливы, бета-аланин, «быстрый эффект», глициновые рецепторы, терморегуляция, клималанин.Rapid effect of beta-alanine in the therapy of hot flashes: a comparative biophysical modeling of interactions beta-alanine, taurine and glycine with the glycine receptors
Torshin I.Y., Gromova O.A., Limanova O.A.Abstract
Beta-alanine is a non-canonical amino acid used in the treatment of hot flashes. In this paper we present the results of biophysical modeling of the interactions of ligands with glycine receptors. It is shown that the binding energy of the receptor beta-alanine greater than the binding energy of the glycine, which corresponds to the prolonged (compared with glycine) effects of beta-alanine on glycine receptors.
Key words: hot flashes, beta-alanine, rapid effects, glycine receptors, thermoregulation, klimalanin.Введение
Cпецифические вегетативно-сосудистые пароксизмы – так называемые приливы – быстро развивающийся вазомоторный симптомокомплекс в перименопаузе. Субъективно «прилив» можно описать как внезапную волну интенсивного тепла и даже жара по телу, сопровождающуюся профузным потоотделением, тахикардией и у многих пациенток подъемом артериального давления.
Препараты для «быстрого» купирования возникающих пароксизом практически неизвестны. Наиболее часто в терапии приливов используется заместительная гормональная терапия (ЗГТ) препаратами эстрогенов. Эффекты синтетических эстрогенов развиваются постепенно, в течение недель и месяцев [1, 2]. Кроме того, пероральная ЗГТ может приводить к головной боли и повышает риск рака молочной железы, яичников, матки, венозной тромбоэмболии [3–6], что простимулировало проведение многочисленных исследований альтернативных видов терапии приливов [7].
Накапливающиеся клинические данные позволяют утверждать, что бета-аланин может купировать приливы в течение нескольких минут. Молекулярно-физиологический механизм развития столь «быстрого» эффекта бетааланина не вполне ясен. Фармакологическое действие бета-аланина объяснимо с точки зрения гормонально-нейротрансмиттерного подхода [8] через воздействие на терморегуляторное ядро в преоптической области гипоталамуса. В терморегуляторном ядре содержатся многочисленные типы рецепторов, которые могут влиять на терморегуляцию: рецепторы гамма-аминомасляной кислоты (ГАМК) [9], гистаминовые [10], альфа-адренергические [11], допаминовые [12], простагландиновые [13, 14], глутаматные, холинергические [15] и глициновые рецепторы [16].
В настоящей работе рассматриваются молекулярно-биофизические механизмы глициновых рецепторов. Интерес к глициновым рецепторам обусловлен тем, что среди вышеперечисленных рецепторов взаимодействие бета-аланина было достоверно доказано только для глициновых рецепторов [8]. Взаимодействуя с глициновыми рецепторами, бета-аланин может способствовать «быстрой» нормализации активности терморегуляторной зоны гипоталамуса, а через синтез пантотената (витамин В5) и накопление карнозина бета-аланин способствует стабилизации энергетического метаболизма, что также соответствует ослаблению симптоматики приливов (рис. 1).
Рис. 1. Механизмы терапевтического воздействия бетааланина при приливах.Заметим, что «медленные» эффекты бета-аланина (днинедели), обусловленные воздействием на энергетический метаболизм (синтез карнозина и пантотената), не менее важны, чем «быстрые» эффекты бета-аланина (минуты), обусловленные взаимодействиями с глициновыми рецепторами (и, возможно, с рецепторами других типов). Стабилизация энергетического метаболизма способствует уменьшению симптоматики приливов [16–19].
Прием бета-аланина постепенно увеличивает уровни карнозина в мышцах, уменьшает утомляемость женщины во время приливов и увеличивает ресурс мышечной системы [20]. В конечном итоге долговременная обеспеченность карнозином нормализует работу гладкой мускулатуры сосудов и тем самым способствует нормализации терморегуляции организма.
Взаимодействие бета-аланина с глициновыми рецепторами является одним из наиболее вероятных объяснений «быстрого» эффекта клималанина. Активированные глициновые рецепторы (глицинуправляемые хлоридные каналы) опосредуют процессы торможения в стволе головного мозга и в спинном мозге. Связываясь с рецептором, нейротрансмиттер глицин и бета-аланин открывают канал рецептора, который пропускает внутрь постсинаптических нейронов ионы хлора. Активация глициновых рецепторов оказывает нормализующее (в случае повышенного нервного возбуждения) действие[21]. В частности, глициновые рецепторы воздействуют на терморегуляторное ядро гипоталамуса [22], активирующее механизмы для поддержания температуры тела в нормальном диапазоне, называемом терморегуляторной зоной. Как известно, у женщин с приливами имеет место сужение границ терморегуляторной зоны [23, 24].
В настоящей работе было проведено сравнительное биофизическое моделирование взаимодействия бета-аланина, таурина и глицина с глициновыми рецепторами. На основе полученных моделей глициновых рецепторов оценивалось сродство (энергия связывания) глицина, бета-аланина и таурина с рецепторами.
Материалы и методы
Модели пространственной структуры глициновых рецепторов человека были приготовлены по методике, описанной в работе [25]. Затем были исследованы возможности размещения молекул лигандов в связывающих карманах рецепторов. Для этого проводилась минимизация энергии при различных начальных расположениях молекулы относительно кармана рецептора [26]; для анализа данных расчетов использовался метод молекулярных энергетических профилей [25], позволяющий анализировать энергетический вклад (dGост) каждого аминокислотного остатка белка.
Результаты и обсуждение
В ходе настоящего исследования были получены различные пространственные структуры глицинового рецептора и рассчитаны значения энергий связывания (отражающих степень сродства к рецептору) рецепторов с глицином, бета-аланином, таурином. Полученные данные были сравнены с имеющимися литературными данными по структурефункции глициновых рецепторов.
Модель пространственной структуры глицинового рецептора
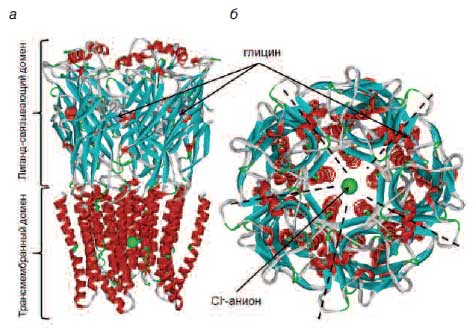
Рецептор глицина является трансмембранным белком, состоящим из пяти белковых субъединиц (рис. 2), образующих центральный канал для селективного проведения ионов хлора [27, 28]. Каждая субъединица состоит из трансмембранного домена (который, собственно, и образует хлоридный канал) и лигандсвязывающего домена (опосредует взаимодействия рецептора с агонистами: глицином, бета-аланином, таурином). К настоящему времени известны 5 типов субъединиц глициновых рецепторов – 4 субъединицы типа альфа (α1, α2, α3, α4) и одна β-субъединица, так что молекула глицинового рецептора может быть образована различными комбинациями этих 5 типов субъединиц.
Рис. 2. Пространственная структура глициновых рецепторов.
Показан ион хлорида в центре канала (зеленая сфера) и участки (участки) связывания молекул глицина (сферические модели): а – Вид сбоку; б – Вид сверху (снаружи нейрона, т.е. со стороны синапса). Штрихпунктирными линиями показано расположение контактов между пятью субъединицами рецептора.Расчет энергий связывания бета-аланина с глициновыми рецепторами
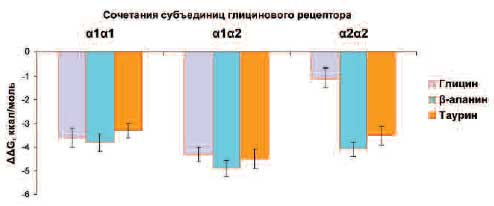
Отметим, что вследствие существования пяти взаимозаменяемых субъединиц (α1, α2, α3, α4 и β), может существовать множество (тысячи) разновидностей глициновых каналов, различающихся как по количественному набору субъединиц (например, 3α1 и 2β и т.д.), так и по порядку включения субъединиц в состав канала (например, α1α2α3α4β, α1α1α2α3β, α1α2α1ββ и десятки других) [28]. Состав субъединиц весьма важен, так как участки связывания глицина и других агонистов расположены в контактах между субъединицами [29], что делает необходимым оценку энергий связывания для разных сочетаний субъединиц (α1α1, α1α2, α2α2 и др.). Так как субъединицы α1 и α2 обнаруживаются наиболее часто [30], в настоящей работе расчет энергий связывания агонистов с глициновыми рецепторами проводился для трех сочетаний субъединиц – α1α1, α1α2 и α2α2 (рис. 3).
Приведенные на рис. 3 значения энергии связывания ΔΔG = ΔGкомп - ΔGсвоб (ΔGкомп – энергия образования комплекса лиганд-рецептор, ΔGсвоб – энергия образования рецептора, не связанного с лигандом) отражают так называемый выигрыш в энергии при образовании комплекса рецептора с глицином, бета-аланином и другими лигандами. Чем более отрицательно значение ΔΔG, тем более прочен комплекс исследуемого лиганда с рецептором (т.е. выше сродство лиганда к рецептору). Как видно из рис. 3, сродство бета-аланина к глициновым рецептором было сравнимо (а в случае α1α2 и α2α2 – даже выше) со сродством самого глицина.
Рис. 3. Энергии связывания глицина, бета-аланина и таурина с различными типами контактов субъединиц рецепторов.
ΔΔG (ккал/моль), энергия связывания лиганда, отражающая сродство лиганда к рецептору: чем более отрицательно значение ΔΔG, тем выше сродство.Молекулярные детерминанты ионной селективности глициновых рецепторов
Аминокислоты-агонисты глициновых рецепторов образуют следующий порядок по степени активации каналов глициновых рецепторов: глицин>бета-аланин>таурин>ГАМК [31–33]. Следует отметить, что определяемая в биохимических экспериментах «степень активации каналов» не отражает «сродства» (т.е. степени связывания) агониста к каналам. Большая энергия связывания (большее сродство) агониста к каналам, даже при более низкой активации, обеспечивает более пролонгированное воздействие агониста на рецептор. Это имеет прямое отношение к результату, полученному в настоящей работе: рассчитанная энергия связывания бета-аланина с глициновым рецептором выше, чем энергия связывания самого глицина с рецептором (или, по крайней мере, сравнима с энергией связывания глицина).
Поэтому клималанин может характеризоваться более продолжительным эффектом воздействия на глициновые рецепторы, чем сам глицин. Подобно глицину бета-аланин в составе клималанина активирует глициновые рецепторы и в коре больших полушарий головного мозга, что в той или иной мере может положительно влиять на когнитивные функции (память и концентрацию внимания). Это открывает еще одну очень интересную перспективу изучения препарата клималанин на когнитивные возможности, как правило, подавленные при патологическом течении периклимактерического периода. Более пролонгированное воздействие бета-аланина на глициновый рецептор при приливах соответствует более продолжительному воздействию препаратов на основе бета-аланина на процессы нормализации работы терморегуляторного ядра. Этот вывод представляется важным для понимания фармакологических эффектов препарата клималанин. Иначе говоря, клималанин может способствовать продолжительной стабилизации функционирования терморегуляторного ядра гипоталамуса. Нами была уточнена структура участка связывания глицина и бета-аланина глициновым рецептором. По данным биохимических исследований, остатки N102, G160, L200, Y202 и T204 (нумерация по последовательности a1 субъединицы) образуют участок связывания агонистов и определяют чувствительность рецептора [34]; лейцин-200 (L200), тирозин-202 (Y202) и треонин-204 (T204) играют особую роль в чувствительности рецептора к глицину [35]. В соответствии с полученными данными, аминокислоты L200, Y202 и T204 поддерживают структуру всего глицинсвязывающего участка (рис. 4). В то же время остатки фенилаланина F159 и тирозина Y161, специфически влияющие на чувствительность к агонистам, расположены в непосредственной близости с молекулами глицина и бета-аланина. Замены аминокислот F159Y и Y161F приводили к увеличению чувствительности к глицину в 12 раз, а чувствительности к бета-аланину – в 120 раз (!) [35].
Рис. 4. Пространственные структуры комплексов 1-субъединицы с глицином и бета-аланином.Следует отметить, что полученные модели комплексов позволяют подробно рассмотреть молекулярные механизмы воздействия ионов цинка и кислотности среды (рН) на активность глициновых рецепторов. Модуляция активности глициновых рецепторов посредством связывания ионов цинка весьма важна для активности рецептора. Как известно, ионы Zn2+ концентрируются в прозрачных пресинаптических пузырьках и секретируются в синаптическую щель одновременно с секрецией соответствующего нейротрансмиттера. Цинк повышает сродство глицина к рецептору, но повышенные концентрации цинка (более 10 мкмоль/л) способствуют снижению сродства [36].
Участок связывания цинка образован аминокислотными остатками H107 и H109, расположенными в лигандсвязывающем домене рецептора (рис. 5). Связывание данными остатками цинка стабилизирует структуру агонистсвязывающего участка, образованного F159 и другими остатками, что и способствует повышению сродства рецептора к агонистам. Избыточные уровни цинка соответствуют связыванию второго иона цинка непосредственно в участке связывания агониста, что и будет приводить к снижению сродства рецептора к глицину и бета-аланину.
Рис. 5. Участки связывания лигандов глицина и бета-аланина на границе между двумя субъединицами типа 1. Показаны молекулы лигандов (сферы темно-зеленого цвета) и аминокислоты, непосредственно участвующие в связывании молекул: а – глицин; б – бета-аланин.Отметим, что у 80–90% женщин в климактерическом периоде развивается дефицит цинка, связанный со снижением кислотности желудка, сокращением микробиоты кишечника и диетарным дефицитом (в частности, вследствие углеводного типа питания). Дефицит цинка является диетарным фактором риска развития инсулинорезистентности, глюкозотолерантности, метаболического синдрома и сахарного диабета [37]. Поэтому эффективность терапии приливов бета-аланином может повышаться при нормальной обеспеченности пациентки цинком. При этом не следует превышать физиологических суточных потребностей в цинке (10–15 мг/сут). Отметим, что в климактерическом периоде дефицит цинка встречается в сотни раз чаще, чем избыточное потребление цинка.
Эффекты кислотности (рН) внеклеточной среды на активность рецепторов обусловлены взаимодействиями протонов с регуляторным цинк связывающим участком рецептора, образованным остатками гистидина H107 и H109 (рис. 6). Чувствительность рецепторов к глицину значительно уменьшается по мере понижения рН спинномозговой жидкости [38].
Рис. 6. Расположения участка связывания цинка в пространственной структуре 1-субъединицы глицинового рецептора.С клинической точки зрения, более низкие уровни рН соответствуют метаболическому ацидозу. Нарушения углеводного обмена способствуют формированию метаболического ацидоза и чаще сопровождаются симптоматикой приливов. Так как при снижении рН сродство лигандов к глициновым рецепторам снижается, для повышения эффективности терапии бета-аланином следует также принять комплекс мер по снижению метаболического ацидоза (снижение потребления углеводов, прием пищевых продуктов, способствующих ощелачиванию крови и др.).
Заключение
Требованием времени является поиск негормональных, но высокоэффективных фармакологических средств для патогенетического лечения приливов. Терапия приливов препаратом клималанин характеризуется «быстрым» эффектом. Одним из наиболее вероятных механизмов «быстрого» воздействия бета-аланина на купирование приливов является активация глициновых рецепторов.
Проведенное в настоящей работе сравнительное моделирование взаимодействий клималанина и глицина с пространственной структурой глициновых рецепторов показало, что энергия связывания рецептором бета-аланина превышает энергию связывания самого глицина. Последнее соответствует пролонгированному воздействию препаратов бета-аланина на глициновые рецепторы, что позволяет предположить, что при применении препарата клималанин возможно улучшение когнитивных функций. Анализ молекулярных детерминант селективности глициновых рецепторов показал, что при планировании терапии приливов бета-аланином целесообразно оценивать наличие у пациентки дефицита цинка и метаболического ацидоза.
Следует подчеркнуть, что способ приема препарата клималанин может оказывать существенное влияние на то, какой эффект будет более выражен – «быстрый» или «медленный». Так, рассасывание таблетки под языком будет способствовать быстрому поступлению бета-аланина в относительно высоких концентрациях непосредственно в кровь со слизистой подъязычного пространства, оказывая воздействие на головной мозг (терморегуляторная зона гипоталамуса) через связывание с глициновыми рецепторами и приводя к «быстрому эффекту» воздействия на терморегуляторное ядро гипоталамуса.
Литература
- Maclennan AH, Broadbent JL, Lester S, Moore V. Oral oestrogen and combined oestrogen/progestogen therapy versus placebo for hot flushes. Cochrane Database Syst Rev 2004; 4: CD002978.
- Reynolds RF, Obermeyer CM, Walker AM, Guilbert D. Side effects and sociobehavioral factors associated with the discontinuation of hormone therapy in a Massachusetts health maintenance organization. Menopause 2001; 8 (3): 189–99.
- Rossouw JE, Anderson GL, Prentice RL et al. Risks and benefits of estrogen plus progestin in healthy postmenopausal women: principal results From the Women's Health Initiative randomized controlled trial. JAMA 2002; 288 (3): 321–33.
- Lacey JV Jr, Mink PJ, Lubin JH et al. Menopausal hormone replacement therapy and risk of ovarian cancer. JAMA 2002; 288 (3): 334–41.
- Coughlin SS, Giustozzi A, Smith SJ, Lee NC. A meta-analysis of estrogen replacement therapy and risk of epithelial ovarian cancer. J Clin Epidemiol 2000; 53 (4): 367–75.
- Tanis BC, van den Bosch MA, Kemmeren JM et al. Oral contraceptives and the risk of myocardial infarction. N Engl J Med 2001; 345 (25): 1787–93.
- Nelson HD, Vesco KK, Haney E et al. Nonhormonal therapies for menopausal hot flashes: systematic review and meta-analysis. JAMA 2006; 295 (17): 2057–71.
- Громова О.А., Торшин И.Ю., Лиманова О.А. Патофизиология вегетативно-сосудистых пароксизмов (приливов) в период менопаузы у женщин и механизмы воздействия бета-аланина. Новая клиникофармакологическая концепция. Вопросы акушерства, гинекологии и перинатологии. 2011; 1.
- Alo R, Avolio E, Di Vito A et al. Distinct alpha subunit variations of the hypothalamic GABAA receptor triplets (alphabetagamma) are linked to hibernating state in hamsters. BMC Neurosci 2010; 11: 111.
- Lundius EG, Sanchez-Alavez M, Ghochani Y et al. Histamine influences body temperature by acting at H1 and H3 receptors on distinct populations of preoptic neurons. J Neurosci 2010; 30 (12): 4369–81.
- Frontini A, Giordano A. Leptin-sensitive neurons in mouse preoptic area express alpha 1A- and alpha 2A-adrenergic receptor isoforms. Neurosci Lett 2010; 471 (2): 83–8.
- Lee TF, Mora F, Myers RD. Dopamine and thermoregulation: an evaluation with special reference to dopaminergic pathways. Neurosci Biobehav Rev 1985; 9 (4): 589–98.
- Nakamura Y, Nakamura K, Morrison SF. Different populations of prostaglandin EP3 receptor-expressing preoptic neurons project to two fever-mediating sympathoexcitatory brain regions. Neuroscience 2009; 161 (2): 614–20.
- Nakamura Y, Nakamura K, Matsumura K et al. Direct pyrogenic input from prostaglandin EP3 receptor-expressing preoptic neurons to the dorsomedial hypothalamus. Eur J Neurosci 2005; 22 (12): 3137–46.
- Takahashi A, Ishimaru H, Ikarashi Y et al. Hypothalamic cholinergic regulation of body temperature and water intake in rats. Auton Neurosci 2001; 94 (1–2): 74–83.
- Karlsson U, Haage D, Johansson S. Currents evoked by GABA and glycine in acutely dissociated neurons from the rat medial preoptic nucleus. Brain Res 1997; 770 (1–2): 256–60.
- Peskov AB, Maevskii EI, Uchitel' ML et al. Succinate-based preparation alleviates manifestations of the climacteric syndrome in women. Bull Exp Biol Med 2005; 140 (3): 312–4.
- Dormire S, Howharn C. The effect of dietary intake on hot flashes in menopausal women. J Obstet Gynecol Neonatal Nurs 2007; 36 (3): 255–62.
- Dormire SL, Reame NK. Menopausal hot flash frequency changes in response to experimental manipulation of blood glucose. Nurs Res 2003; 52 (5): 338–43.
- Hill CA, Harris RC, Kim HJ et al. Influence of beta-alanine supplementation on skeletal muscle carnosine concentrations and high intensity cycling capacity. Amino Acids 2007; 32 (2): 225–33.
- Lynch JW. Native glycine receptor subtypes and their physiological roles. Neuropharmacology 2009; 56 (1): 303–9.
- Casper RF, Yen SS. Neuroendocrinology of menopausal flushes: an hypothesis of flush mechanism. Clin Endocrinol (Oxf) 1985; 22 (3): 293–312.
- Freedman RR, Krell W. Reduced thermoregulatory null zone in postmenopausal women with hot flashes. Am J Obstet Gynecol 1999; 181 (1): 66–70.
- Harris RC, Tallon MJ, Dunnett M et al. The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids 2006; 30 (3): 279–89.
- Torshin IY. Bioinformatics in the Post-Genomic Era: The Role of Biophysics 2006 Nova Biomedical Books, NY, ISBN: 1-60021-048, p. 96–108.
- Торшин И.Ю., Громова О.А., Рудаков К.В. Сравнительный анализ дидрогестерона и других прогестагенов: расчет энергий связывания со стероидными рецепторами. Вопросы гинекологии, акушерства и перинатологии. 2011; 2 (10): 68–80.
- Lynch JW. Molecular structure and function of the glycine receptor chloride channel. Physiol Rev 2004; 84 (4): 1051–95.
- Langosch D, Thomas L. Conserved quaternary structure of ligand-gated ion channels: the postsynaptic glycine receptor is a pentamer. Proc Natl Acad Sci USA1988; 85: 7394–8.
- Corringer PJ, Le Novere N, Changeux JP. Nicotinic receptors at the amino acid level. Annu Rev Pharmacol Toxicol 2000; 40: 431–58.
- Mohammadi B, Krampfl K, Cetinkaya C et al. Kinetic analysis of recombinant mammalian -1 and -1 -glycine receptor channels. Eur Biophys J 2003; 32: 529–36.
- Lester HA. The permeation pathway of neurotransmitter-gated ion channels. Annu Rev Biophys Biomol Struct 21: 267–292, 1992.
- Bormann J, Rundstrom N, Betz H. Residues within transmembrane segment M2 determine chloride conductance of glycine receptor homo- and hetero-oligomers. EMBO J 1993; 12: 3729–37.
- Lynch JW, Rajendra S, Pierce KD et al. Identification of intracellular and extracellular domains mediating signal transduction in the inhibitory glycine receptor chloride channel. EMBO J 1997; 16: 110–20.
- Han NL, Haddrill JL, Lynch JW. Characterization of a glycine receptor domain that controls the binding and gating mechanisms of the в-amino acid agonist, taurine. J Neurochem 2001; 79: 636–47.
- Rajendra S, Vandenberg RJ, Pierce KD et al. The unique extracellular disulfide loop of the glycine receptor is a principal ligand binding element. EMBO J 1995; 14: 2987–98.
- Laube B, Kuhse J, Rundstrom N et al. Modulation by zinc ions of native rat and recombinant human inhibitory glycine receptors. J Physiol 1995; 483: 613–9.
- Ребров ВГ, Громова ОА. Витамины, макро- и микроэлементы, ГЭОТАР-МЕД. 2008; с. 647.
- Chen Z, Dillon GH, Huang R. Molecular determinants of proton modulation of glycine receptors. J Biol Chem 2004; 279: 876–83.
| Апрель 2012 г. |